Къде се намират рецепторите за болка при хората? Рецептори за болка: местоположение, особености на нервната система
Рецепторите за болка (ноцицептори) реагират на стимули, които заплашват тялото с увреждане. Има два основни типа ноцицептори: Adelta механоноцицептори и полимодални C ноцицептори (има няколко други типа). Както подсказва името им, механоноцицепторите се инервират от тънки миелинизирани влакна, а полимодалните С-ноцицептори се инервират от немиелинизирани С-влакна. Делта-механоноцицепторите реагират на силно механично дразнене на кожата, например убождане с игла или щипка с пинсети. Те обикновено не реагират на термични и химични болезнени стимули, освен ако преди това не са били сенсибилизирани. Обратно, мултимодалните С-ноцицептори реагират на болкови стимули от различен тип: механични, температурни (фиг. 34.4) и химични.
В продължение на много години не беше ясно дали болката е резултат от активирането на специфични влакна или от свръхактивност на сензорни влакна, които обикновено имат други модалности. Последната възможност изглежда е по-съвместима с обичайния ни опит. С възможното изключение на обонянието, всеки сензорен стимул с прекомерна интензивност - заслепяваща светлина, пронизващ ушите звук, силен удар, топлина или студ извън нормалните граници - води до болка. Тази гледна точка на здравия разум е изразена от Еразъм Дарвин в края на 18 век и Уилям Джеймс в края на 19 век. Здравият разум обаче тук (както и навсякъде) оставя какво да се желае. Понастоящем няма съмнение, че в повечето случаи усещането за болка възниква в резултат на стимулиране на специализирани ноцицептивни влакна. Ноцицептивните влакна нямат специализирани окончания. Те присъстват под формата на свободни нервни окончания в дермата на кожата и на други места в тялото. Хистологично те са неразличими от С-механорецепторите (МЕХАНОСЕНЗИТИВНОСТ) и - и А-делта терморецепторите (глава ТЕРМИЧНА ЧУВСТВИТЕЛНОСТ). Те се различават от споменатите рецептори по това, че прагът на адекватните им стимули е по-висок от нормалния диапазон. Те могат да бъдат разделени на няколко различни типа въз основа на критерия коя сетивна модалност осигурява адекватен стимул за тях. Вредни термични и механични стимули се откриват от миелинизирани влакна с малък диаметър, Таблица 2.2 показва, че те са класифицирани като делта влакна от категория А. Полимодалните влакна, които реагират на голямо разнообразие от интензитети на стимули с различни модалности, също са с малък диаметър, но не са миелинизирани. Таблица 2.2 показва, че тези влакна са клас C. Делта влакната провеждат импулси с честота 5-30 m / s и са отговорни за "бърза" болка, остро усещане за пробождане; С-влакната провеждат по-бавно - 0,5 - 2 m/s и сигнализират за "бавна" болка, често продължителна и често преминаваща в тъпа болка. AMT (механо-термо-ноцицептори с A делта влакна) се разделят на два типа. AMT тип 1 се среща главно в неокосмената кожа. AMT тип 2 се намират главно в окосмената кожа.Накрая, ноцицепторите на С-влакна (CMT влакна) имат праг в диапазона от 38°C - 50°C и реагират с постоянна активност, която зависи от интензивността на стимула (фиг. 21.1а). AMT и CMT рецепторите, както показват имената им, реагират както на термични, така и на механични стимули. Физиологичната ситуация обаче далеч не е проста. Механизмът на предаване на тези два модалности е различен. Прилагането на капсаицин не повлиява чувствителността към механични стимули, но инхибира реакцията към термични. Освен това, докато капсаицинът има аналгетичен ефект върху термичната и химическа чувствителност на мултимодални С-влакна в роговицата, той не засяга механочувствителността. И накрая, доказано е, че механичните стимули, които генерират същото ниво на активност в SMT влакната като термичните, въпреки това причиняват по-малко болка. Може би неизбежно по-широката повърхностна площ, покрита от термичен стимул, включва активността на повече CMT влакна, отколкото би бил случаят с механичен стимул.
Сенсибилизацията на ноцицепторите (повишена чувствителност на аферентните рецепторни влакна) възниква след отговора им на вредно дразнене. Сенсибилизираните ноцицептори реагират по-интензивно на повтарящ се стимул, тъй като техният праг е понижен (фиг. 34.4). В този случай се наблюдава хипералгезия - по-силна болка в отговор на стимул със същата интензивност, както и намаляване на прага на болката. Понякога ноцицепторите генерират фонов разряд, който причинява спонтанна болка.
Сенсибилизация възниква, когато химични фактори като K+ йони, брадикинин, серотонин, хистамин, ейкозаноиди (простагландини и левкотриени) се отделят близо до ноцицептивните нервни окончания в резултат на тъканно увреждане или възпаление. Да кажем, че вреден стимул удря кожата и унищожава клетките на тъканната област близо до ноцицептора (фиг. 34.5, а). K+ йони излизат от умиращите клетки, които деполяризират ноцицептора. Освен това се освобождават протеолитични ензими; когато взаимодействат с глобулините на кръвната плазма, се образува брадикинин. Той се свързва с рецепторните молекули на ноцицепторната мембрана и активира втората система за съобщения, която сенсибилизира нервното окончание. Други освободени химикали, като тромбоцитен серотонин, мастоцитен хистамин и ейкозаноиди на различни клетъчни елементи, допринасят за сенсибилизация чрез отваряне на йонни канали или активиране на вторични системи за съобщения. Много от тях също засягат кръвоносните съдове, клетките на имунната система, тромбоцитите и други ефектори, участващи във възпалението.
В допълнение, активирането на ноцицепторния терминал може да освободи регулаторни пептиди като субстанция P (SP) и калцитонин ген-кодиран пептид (CGRP) от други терминали на същия ноцицептор чрез аксоновия рефлекс (фиг. 34.5b). Нервен импулс, възникващ в един от клоновете на ноцицептора, се насочва по майчиния аксон към центъра. В същото време той се разпространява антидромно по периферните клонове на аксона на същия ноцицептор, което води до освобождаване на вещество Р и CGRP в кожата (фиг. 34.5, b). Тези пептиди причиняват
Болката е симптом на много заболявания и наранявания на тялото. Човек е разработил сложен механизъм за възприемане на болка, който сигнализира за увреждане и принуждава човек да предприеме мерки за отстраняване на причините за болка (дърпане на ръката и др.).
Ноцицептивна система
Така нареченият ноцицептивна система. В опростена форма механизмът на болката може да бъде представен по следния начин (Фигура ⭣).
При дразнене на болкови рецептори (ноцицептори), локализирани в различни органи и тъкани (кожа, кръвоносни съдове, скелетни мускули, периост и др.), възниква поток от болкови импулси, които преминават през аферентни влакна към дорзалните рога на гръбначния мозък. .
Аферентните влакна са два вида: А-делта влакна и С-влакна.
А-делта влакноса миелинизирани, което означава, че са бързопроводими - скоростта на импулсите през тях е 6-30 m/s. А-делта влакната са отговорни за предаването на остра болка. Те се възбуждат от силно интензивно механично (убождане с игла), а понякога и термично дразнене на кожата. Те по-скоро имат информационна стойност за тялото (принуждават ви да дръпнете ръката си, да отскочите и т.н.).
Анатомично А-делта ноцицепторите са представени от свободни нервни окончания, разклонени под формата на дърво. Те се намират предимно в кожата и в двата края на храносмилателния тракт. Те се намират и в ставите. Предавателят (предавател на нервни сигнали) на А-делта влакна остава неизвестен.
С-влакна- немиелинизирани; те провеждат мощни, но бавни импулсни потоци със скорост 0,5-2 m/s. Смята се, че тези аферентни влакна са посветени на възприемането на вторична остра и хронична болка.
С-влакната са представени от плътни, некапсулирани гломерулни тела. Те са полимодални ноцицептори, следователно реагират както на механични, така и на термични и химични стимули. Те се активират от химикали, които се появяват по време на увреждане на тъканите, като в същото време са хеморецептори, те се считат за оптимални рецептори за увреждане на тъканите.
С-влакната се разпространяват във всички тъкани с изключение на централната нервна система. Влакната, които имат рецептори, които усещат увреждане на тъканите, съдържат вещество P, което действа като предавател.
В дорзалните рога на гръбначния мозък сигналът се превключва от аферентното влакно към интерневрона, от който на свой ред импулсът се разклонява, вълнувайки моторните неврони. Това разклонение е придружено от двигателна реакция на болка - отдръпване на ръката, отскачане и др. От интернейрона потокът от импулси, издигайки се по-нататък през централната нервна система, преминава през продълговатия мозък, който съдържа няколко жизненоважни центъра: дихателни, вазомоторни, центрове на блуждаещия нерв, център за кашлица, център за повръщане. Ето защо болката в някои случаи има вегетативен придружител - сърцебиене, изпотяване, скокове на кръвното налягане, слюноотделяне и др.
След това болковият импулс достига таламуса. Таламусът е една от ключовите връзки в предаването на сигнала за болка. Той съдържа така наречените превключващи (SNT) и асоциативни ядра на таламуса (AT). Тези образувания имат определен, доста висок праг на възбуждане, който не всички болкови импулси могат да преодолеят. Наличието на такъв праг е много важно в механизма на възприемане на болката, без него всяко най-малко дразнене би причинило болезнено усещане.
Но ако импулсът е достатъчно силен, той предизвиква деполяризация на PAT клетките, импулси от тях навлизат в двигателните зони на кората на главния мозък, определяйки самото усещане за болка. Този път на болковите импулси се нарича специфичен. Осигурява функция за сигнализиране на болка - тялото възприема появата на болка.
На свой ред, активирането на AYT предизвиква импулси да навлязат в лимбичната система и хипоталамуса, осигурявайки емоционално оцветяване на болката (неспецифичен път на болка). Именно поради този път възприемането на болката има психо-емоционална конотация. Освен това, благодарение на този път, хората могат да опишат възприеманата болка: остра, пулсираща, пронизваща, боляща и т.н., което се определя от нивото на въображение и вида на нервната система на човека.
Антиноцицептивна система
В цялата ноцицептивна система има елементи на антиноцицептивната система, която също е неразделна част от механизма на възприемане на болката. Елементите на тази система са предназначени да потискат болката. Механизмите на развитие на аналгезия, контролирани от антиноцицептивната система, включват серотонинергичната, GABAergic и в най-голяма степен опиоидната система. Функционирането на последните се осъществява благодарение на протеинови предаватели - енкефалини, ендорфини - и специфични за тях опиоидни рецептори.
Енкефапини(мет-енкефалин - H-Tyr-Gly-Gly-Phe-Met-OH, лев-енкефалин - H-Tyr-Gly-Gly-Phe-Leu-OH и др.) са изолирани за първи път през 1975 г. от мозъка на бозайници . Според химичната си структура те принадлежат към класа на пентапептидите, имащи много сходна структура и молекулно тегло. Енкефалините са невротрансмитери на опиоидната система, функциониращи по цялата й дължина от ноцицепторите и аферентните влакна до мозъчните структури.
Ендорфини(β-ендофин и динорфин) са хормони, произвеждани от кортикотропни клетки на средния дял на хипофизната жлеза. Ендорфините имат по-сложна структура и по-голямо молекулно тегло от енкефалините. По този начин β-ендофинът се синтезира от β-липотропин, като всъщност е част от 61-91 аминокиселина на този хормон.
Енкефалините и ендорфините, стимулиращи опиоидните рецептори, извършват физиологична антиноцицепция и енкефалините трябва да се разглеждат като невротрансмитери, а ендорфините като хормони.
Опиоидни рецептори- клас рецептори, които, като мишени за ендорфини и енкефалини, участват в осъществяването на ефектите на антиноцицептивната система. Името им идва от опиум - изсушен млечен сок от сънотворния мак, известен от древността като източник на наркотични аналгетици.
Има 3 основни типа опиоидни рецептори: μ (mu), δ (делта), κ (kappa). Тяхната локализация и ефектите, които възникват при възбуждането им, са представени в таблица ⭣.
| Локализация | Ефект при възбуда | |
| μ рецептори: | ||
| Антиноцицептивна система | ➔ | Аналгезия (спинална, супраспинална), еуфория, пристрастяване. |
| Cortex | ➔ | Кортикално инхибиране, сънливост. Индиректно - брадикардия, миоза. |
| Дихателен център | ➔ | Респираторна депресия. |
| Център за кашлица | ➔ | Потискане на кашличния рефлекс. |
| Център за повръщане | ➔ | Стимулиране на центъра за повръщане. |
| Хипоталамус | ➔ | Инхибиране на центъра за терморегулация. |
| хипофиза | ➔ | Отслабване на производството на гонадотропни хормони и увеличаване на производството на пролактин и антидиуретичен хормон. |
| Стомашно-чревния тракт | ➔ | Намалена перисталтика, спазъм на сфинктера, отслабена секреция на жлезите. |
| δ рецептори: | ||
| Антиноцицептивна система | ➔ | Аналгезия. |
| Дихателен център | ➔ | Респираторна депресия. |
| κ рецептори: | ||
| Антиноцицептивна система | ➔ | Аналгезия, дисфория. |
Енкефалините и ендорфините, стимулиращи опиоидните рецептори, предизвикват активиране на G₁ протеина, свързан с тези рецептори. Този протеин инхибира ензима аденилат циклаза, който при нормални условия насърчава синтеза на цикличен аденозин монофосфат (цАМР). На фона на неговата блокада, количеството на сАМР вътре в клетката намалява, което води до активиране на мембранните калиеви канали и блокада на калциевите канали.
Както знаете, калият е вътреклетъчен йон, калцият е извънклетъчен йон. Тези промени във функционирането на йонните канали причиняват освобождаването на калиеви йони от клетката, докато калцият не може да навлезе в клетката. В резултат зарядът на мембраната рязко намалява и се развива хиперполяризация - състояние, при което клетката не възприема и не предава възбуждане. В резултат на това възниква потискане на ноцицептивните импулси.
източници:
1. Лекции по фармакология за висше медицинско и фармацевтично образование / V.M. Брюханов, Я.Ф. Зверев, В.В. Лампатов, А.Ю. Жариков, О.С. Талалаева - Барнаул: Издателство "Спектр", 2014 г.
2. Обща човешка патология / Саркисов Д.С., Палцев М.А., Хитров Н.К. - М.: Медицина, 1997.
|
Ярослав Алексеевич Андреев- Кандидат на биологичните науки, старши изследовател в Лабораторията по неврорецептори и неврорегулатори, Департамент по молекулярна невробиология, Институт по биоорганична химия на име. Академици М. М. Шемякин и Ю. А. Овчинников РАН. Научните интереси са свързани с търсене и характеризиране на модулатори на болкови рецептори. |
|
Юлия Александровна Логашина- младши научен сътрудник в същата лаборатория. Той се занимава с търсене и характеризиране на нови TRPA1 рецепторни лиганди. |
|
Ксения Игоревна Любова- студент на Биологическия факултет на Московския държавен университет. М. В. Ломоносов. Изследва TRP рецепторите и техните модулатори. |
|
Александър Александрович Василевски- Кандидат на химическите науки, ръководител на групата от молекулярни инструменти за невробиология, Департамент по молекулярна невробиология, Институт по биоорганична химия на името на. Академици М. М. Шемякин и Ю. А. Овчинников РАН. Специалист в областта на йонните канали и естествените токсини. |
|
Сергей Александрович Козлов- Доктор на химическите науки, ръководител на лабораторията по неврорецептори и неврорегулатори на същия отдел. Област на научни интереси: протеинови рецептори в нервната система и техните лиганди. |
Казват, че животът е болка. Въпреки че тази фраза съдържа нещо негативно, свързано с неприятни усещания, преживявания или дори тежко страдание, не бива да забравяме, че болката (ноцицепцията) ни предупреждава за опасност – тя сигнализира за смущения в организма, който веднага започва да ги премахва. В същото време има и болка, която носи само мъка.
Основната причина за появата на такава болка са неуспехи в предаването на сигнали за болка (нервни импулси) от чувствителни неврони към мозъка, което създава неприятни усещания. Когато излагането на невредни стимули се прецени от невроните за разпознаване като вредно, се развива състояние, наречено свръхчувствителност. И това не винаги е лошо, тъй като в точното време играе важна роля в процеса на оздравяване и възстановяване на тялото. Случва се обаче и да няма истинска причина и свръхчувствителността води до изтощителна хронична болка. В този случай най-често срещаните безвредни стимули (леко докосване или топлина) причиняват алодиния (от гръцки άλλος - друг и οδύνη - мъчение), а болезнените стимули причиняват болка с още по-голяма интензивност, хипералгезия (от гръцки ὑπέρ - над-и ἄλγος - болка). Често необичайно интензивна и често хронична болка, която е инвалидизираща както физиологично, така и психологически, а също така усложнява възстановяването, възниква в резултат на заболявания като артрит, херпес зостер, СПИН, рак на костите и др.
Преди да обвиняваме сетивните неврони (ноцицептори), които възприемат, анализират и предават сигнали за болка, за аномалии, нека разберем как работят в здраво тяло и какво се случва при патологии.
Защо боли толкова много?
Биологичната функция на ноцицепторите е не само да регистрират стимул и да го докладват на мозъка ни, но и да възприемат сигнали от близките съседи. Невроните са заобиколени от други клетки на тялото и междуклетъчната среда, за безопасността и правилното функциониране на които отговаря нашата нервна система. Следователно ноцицепторите имат много молекулярни сензори (или рецептори), конфигурирани да разпознават химични стимули, промени в състава и свойствата на междуклетъчната среда и освобождаването на сигнални молекули от близките клетки. Невронът независимо „изчислява“ приноса на всеки такъв молекулярен сензор въз основа на силата и продължителността на стимулацията и ако стимулите се считат за нежелани, той сигнализира това - и ние чувстваме болка; това е "нормална" физиологична болка (ноцицепция). Патологичната болка възниква както в случай на смърт на неврони поради увреждане на проводимата мрежа на периферната или централната нервна система, така и когато самите неврони работят погрешно и правят грешки поради неизправност на техните сензори.
Сензорите за болка (или рецептори) са мембранни протеини, които разпознават физически или химични стимули върху невронната мембрана. Освен това те са катион-селективни йонни канали, т.е. осигуряват преминаването на положително заредени йони (натрий, калий, калций) през клетъчната мембрана. Активирането на рецепторите води до отваряне на катионни канали и възбуждане на чувствителни неврони - възникване на нервен импулс. По-долу ще ви разкажем повече за най-изследваните рецептори за болка.
Какво се случва, ако предположим, че човек по невнимание изгори ръката си с горещ предмет? Този опасен температурен ефект се регистрира от рецептори, разположени в ноцицепторната мембрана. Те моментално разпознават силната стимулация и предават импулса към централната нервна система. Мозъкът веднага реагира на такова силно вълнение и ние рефлексивно отдръпваме ръката си от горещия предмет. Интересното е, че същите сензори реагират на капсаицин, активното вещество в лютите чушки, което предизвиква „пожар“ в устата.
Други рецептори са отговорни за разпознаването на редица опасни химични ефекти, които възприемат стимули само от вътреклетъчната страна, следователно, за да ги активират, опасните вещества трябва не само да проникнат през кожата, но и да попаднат вътре в неврона, „проправяйки си път“ през липидната биомембрана. Ако химическото изгаряне е причинено от киселина, тогава рецепторът, който е чувствителен към промените в киселинността на околната среда, ще работи и също ще даде силен отговор веднага щом киселината достигне неврона.
Издърпахме ръката си, но по време на контакт с горещата повърхност някои от нашите клетки умряха и в отговор на увреждане на тъканите започва да се развива възпалителен процес. В това участва и нашата нервна система. От увредените клетки, чрез разкъсани цитоплазмени мембрани, молекули, характерни за вътреклетъчната среда, по-специално аденозинтрифосфорна киселина (АТФ), започват да се освобождават в извънклетъчната среда. В този случай невроните също имат собствен рецептор, който се активира от молекулите на АТФ и сигнализира, че в близост до него е настъпила клетъчна смърт и е необходимо тяхното възстановяване. Факт е, че АТФ, както се знае от училище, е основната енергийна молекула на тялото и такава „ценност“ рядко попада в междуклетъчната среда.
Невронът не само сигнализира, той освобождава специални биологично активни съединения, медиатори на възпалението, в извънклетъчната среда, което води до дългосрочно развитие на неврогенно възпаление - вазодилатация и привличане на клетките на имунната система. Докато процесът на регенерация е в ход и в околната среда има медиатори на възпалението, сензорните неврони изпращат сигнал до централната нервна система, където той също се възприема като болка, но не толкова силна. Тъй като увредената тъкан се нуждае от защита, чувствителността на невроните към външни влияния се увеличава и дори незначителни механични или термични ефекти ще предизвикат силна реакция на болка. Това е „благоприятна“ свръхчувствителност.
Почти всеки знае, че се препоръчва да се прилага студено върху увредената тъкан, за да се облекчи болката и да се намали възпалението. Невронните рецептори също участват в този ефект. Основният „студен“ рецептор - ментол (помните ли „ментовия“ студ?) - не се намира в същите неврони, където се намира „топлинният“ и следователно усещанията за студ и топлина се предават от различни сензорни влакна. Оказва се, че информацията от различни ноцицептори се „сумира“ в гръбначния мозък, сигналът от горещото въздействие се коригира, като се вземе предвид сигналът от студа и затова приложеното парче лед може да облекчи силната болка.
Описаната схема за развитие на болката е значително опростена (фиг. 1). Всъщност, за да разберат детайлите на ноцицепцията, учените изучават всеки рецептор поотделно в изолирани условия. Провеждат се експерименти върху клетъчни линии, в които се вмъкват гени за определени рецептори с помощта на методи на генно инженерство. Нека поговорим малко за изследването и функциите на няколко от най-важните рецептори за болка. Както се оказа, те не винаги са фокусирани върху разпознаването и генерирането на сигнал за болка, но участват в регулирането на много други процеси, така че способността да се коригира работата им с различни лекарства ще помогне за лечението на различни заболявания (фиг. 2 ).

Рецептори за температурни и химични стимули
Много често сетивните неврони, които са отговорни за възприемането на висока температура, играят роля в развитието на болка и възпаление. Още в средата на 20-ти век е открито, че големи дози капсаицин предизвикват нов тип обезболяване (аналгезия) при опитни животни. След прилагане на капсаицин първоначално се наблюдава характерна поведенческа реакция, причинена от болка, но след това настъпва дълъг период на загуба на чувствителност към редица външни стимули. Животните в това състояние реагират нормално на лека механична стимулация, но губят реакцията си на много болезнени стимули и не развиват неврогенно възпаление. По този начин невроните, отговорни за усещането на топлина, са отговорни и за усещането на химически стимули и неврогенния компонент на възпалителния отговор. Стана ясно, че рецепторът, който реагира на температурата и капсаицина, може да бъде полезна цел при търсенето на лечения за възпаление и болка. В края на ХХ век. Този рецептор е характеризиран на молекулярно ниво и е наречен TRPV1. преходен рецептор потенциален канал ванилоиден член на семейството 1- първият представител на семейството на ванилоидни рецептори с променлив рецепторен потенциал), или по-просто - ванилоиден рецептор 1 (фиг. 3). Името "ванилоидни рецептори" не е дадено случайно: TRPV1 и други членове на семейството се активират от химични съединения, съдържащи ванилинова група (например капсаицин). Установено е, че TRPV1 е катион-селективен йонен канал, който се активира от различни стимули (температури над 43°C, ниско pH, капсаицин), като освен това неговата активност се регулира от възпалителни медиатори, макар и не директно, а чрез вътреклетъчни посредници. TRPV1 нокаут мишки (т.е. тези, при които генът за този рецептор липсва или е повреден, така че да не работи) реагират много по-бавно на топлина и показват малка или никаква термична свръхчувствителност по време на възпаление. TRPV1 играе важна роля при редица патологични състояния: болка, причинена от възпаление, рак, невропатична и висцерална болка, както и респираторни заболявания, панкреатит и мигрена.

Изследването на TRPV1 доведе до интензивно изследване на подобни рецептори. Така е открит друг ванилоиден рецептор - TRPV3. Интересното е, че той реагира както на приятна топлина, така и на болезнена топлина: активността на TRPV3 се записва при температури над 33°C, като реакцията му е по-силна при по-високи температури и се увеличава при повтаряща се термична стимулация. Освен от температурата, този рецептор се активира и от камфора, лютивите екстракти от мащерка, риган и карамфил. TRPV3 е друг кандидат за свръхчувствителност към болка и неговата активност се регулира от възпалителни медиатори. И накрая, той се активира директно от азотен оксид II (NO), втори носител, който повишава чувствителността на невроните към стимулация. Също така забележително е присъствието на TRPV3 в клетките на кератиноцитната кожа, където неговото активиране води до освобождаване на възпалителния медиатор интерлевкин-1, подчертавайки важната роля на този рецептор при възпалителни кожни заболявания.
TRP рецепторите са тетрамери (фиг. 3), т.е. те са образувани от четири полипептидни вериги. В този случай, както хомомерите, т.е. рецепторите, образувани от идентични вериги (например, TRPV1 или TRPV3, описани по-горе), така и хетеромерите от различни вериги могат да бъдат сглобени. Хетеромерните рецептори (например, изградени от вериги TRPV1 и TRPV3) имат различна чувствителност към топлинни стимули; праговата температура за тяхното активиране е между праговите стойности за хомомерните рецептори.
Интересна история е откриването на рецептора за студ TRPM8 (тук „М“ означава меластатин, което показва функцията на рецепторите от това семейство в меланоцитите, клетките на кожата, отговорни за пигментацията). Първоначално е открит генът, който го кодира, чиято активност се повишава при рак на простатата и някои други онкологични заболявания. Много по-късно беше показано, че TRPM8 реагира на ментол (компонент на мента) и редица други „освежаващи” вещества, както и на понижаване на температурата (под 26°C). Този рецептор сега се счита за основния сензор за студ в нервната система. Изследванията са установили, че TRPM8 е отговорен за широк диапазон на възприемане на студени стимули - от приятна прохлада до болезнен студ и свръхчувствителност към студ. Това разнообразие от функции се обяснява със съществуването на няколко субпопулации от сензорни неврони, които използват TRPM8 като мултифункционален сензор за студ, настроен на определена температура с участието на вътреклетъчни сигнални системи.
Най-неразбираемият и много важен рецептор TRPA1 (тук "А" означава "анкирин", което показва наличието в структурата на рецепторите от това семейство на голям брой "анкиринови повторения", специални протеинови елементи) се намира в сензорните неврони на кожата, епителните клетки на червата, белите дробове и пикочните пътища, пикочния мехур, като TRPA1 често е в съседство с TRPV1. Веществата, които активират TRPA1, причиняват парене, механична и термична свръхчувствителност и неврогенно възпаление. Свръхекспресията на гена, кодиращ TRPA1, води до хроничен кожен сърбеж и алергичен дерматит. Синдромът на наследствено заболяване епизодична болка, което се характеризира с внезапна поява на изтощителна болка по време на гладуване или упражнения, е свързано с мутация в този рецептор, което води до неговата прекомерна активност.
Основната функция на TRPA1 е разпознаването на химически и възпалителни агенти, като техният обхват е толкова голям, че почти всички жизнени процеси на нашето тяло са свързани с правилното функциониране на този рецептор. В дихателната система той разпознава летливи вредни вещества: сълзотворен газ, озон, алдехиди (акролеин, компоненти на канелата), органосерни съединения (остри компоненти на горчица, лук и чесън), причиняващи кашлица, кихане и образуване на слуз. В червата TRPA1 открива наличието на възпалителни агенти. Свръхактивният пикочен мехур при диабет се причинява от активиране на този рецептор от акролеин, който се натрупва в урината. TRPA1 е замесен в развитието на мигрена под въздействието на цигарен дим и формалдехид при някои хора.
Насочването на лекарствата към сензорните невронни рецептори, участващи във възприемането на температурата, води до облекчаване на болката и възпалението. Ето как, без да знае за молекулярните цели, традиционната медицина в различни времена е използвала тинктури от пипер (TRPV1), горчица (TRPA1), мента (TRPM8) и карамфил (TRPV3) за лечение на редица възпалителни заболявания.
Пуринови рецептори
Вече споменахме, че е много важно тялото да знае за увреждането на тъканите. По време на наранявания, когато целостта на органите е нарушена и настъпва клетъчна смърт, молекулите на АТФ навлизат в междуклетъчното пространство по време на исхемия или възпаление. Този коензим на много реакции осигурява енергия за много процеси в клетката; той е твърде ценен за функционирането на клетките, затова рядко се изхвърля от тях. Възприемането на повишаване на локалната концентрация на АТФ се осъществява от пуринергични рецептори (P2X), които са катион-селективни йонни канали; те предизвикват реакция на болка, която възниква поради разрушаване на тъканите, деформация на органи и развитие на тумор. Сензорните неврони се характеризират с подтипове P2X2 и P2X3, като важната роля на последния в развитието на болка по време на възпаление е показана в проучвания върху нокаут мишки. Известно е също, че P2X рецепторите са от фундаментално значение за много физиологични процеси, като регулиране на съдовия тонус, вкусово възприятие и др.
Киселинни рецептори
За да открият киселинността, много видове клетки в нервната система съдържат така наречените киселинно-чувствителни йонни канали ( киселинночувствителни йонни канали,ASIC). Смята се, че те медиират сигнал, свързан с локални промени в pH по време на нормална невронна активност в централната нервна система. Те обаче участват и в патологични процеси. Напоследък рецепторът от подтип ASIC1a се счита за един от основните фактори за смъртта на неврони в централната нервна система по време на исхемични състояния. По време на исхемия и хипоксия гликолизата се увеличава, което води до натрупване на млечна киселина и последващо "подкисляване" на тъканта. "Изключването" на ASIC1a рецептора причинява невропротективни ефекти в исхемичен модел, както е показано при нокаут мишки. В периферната нервна система и висцералните тъкани ASIC са отговорни за чувствителността към болка в резултат на тъканна ацидоза в мускулите, сърдечна исхемия, увреждане на роговицата, възпаление, неоплазми и локална инфекция. Невроните на периферната нервна система съдържат главно рецептори от подтипа ASIC3, чиято активност също трябва да бъде намалена, за да се облекчи болката.
За разлика от TRP рецепторите, P2X рецепторите и ASIC са тримери (фиг. 3), т.е. съставени от три полипептидни вериги. Но по същия начин тези рецептори могат да бъдат хомомери и хетеромери, което увеличава тяхното разнообразие и обхват от функции.
Как да преодолеем болката?
И така, какво трябва да направим, ако изпитваме болка? Независимо дали болката е остра или хронична, тя не може да бъде толерирана и трябва да се използват болкоуспокояващи, за да се върне нашата ноцицептивна система към нормалното и ние самите да живеем в истинския смисъл на думата. В момента много лекарства от различни фармакологични групи се използват за облекчаване на болката. Основното място в тази серия заемат нестероидните противовъзпалителни средства (НСПВС), антиконвулсантите и антидепресантите, както и наркотичните аналгетици (морфин и други опиати и опиоиди). Наличните в момента аналгетици влияят главно на предаването и разпространението на болката. На пазара няма лекарства за специфичната регулация на рецепторите за болка, описани по-горе.
Първата цел за „болка“ за фармацевтичните компании беше рецепторът TRPV1, тъй като сензорните неврони, които го съдържат, играят ролята на интегратори на много стимули, възприемани като болка. Скринингът на химическите библиотеки и рационалното проектиране на лиганди, базирани на познаването на мястото на свързване на капсаицин, доведоха до разработването на значителен брой високоефективни малки молекулни инхибитори на TRPV1. Тези съединения имат аналгетичен ефект, но водят до развитие на хипертермия - повишаване на телесната температура (с 1,5-3 ° C). Хипертермията се превърна в основната причина за отказа на фармацевтичните компании да разработят лекарства, базирани на пълни антагонисти на рецептора TRPV1. Въпреки това, ако този рецептор е само частично инхибиран, повишаването на телесната температура може да бъде избегнато. И ние, под ръководството на академик Е. В. Гришин (1946–2016), успяхме да намерим такива частични инхибитори на TRPV1 в отровата на морски анемони Heteractis crispa. Три пептида бяха открити в отровата на анемона, които инхибират TRPV1 и не повишават телесната температура [, ], но пептидът, наречен ARHC3, имаше най-мекия ефект. Има силен аналгетичен ефект в дози от 0,01–0,1 mg/kg телесно тегло и слабо понижава телесната температура (само с 0,6°C). По отношение на облекчаване на болката той е сравним с морфина, но не предизвиква наркотични ефекти или пристрастяване. Според предклиничните проучвания пептидът е напълно подходящ за по-нататъшни клинични изпитвания, тъй като не са открити странични ефекти върху лабораторни животни. Освен това, понижаването на телесната температура е необходимо, например, за осигуряване на невропротекция при оцелели от сърдечен арест, а хипотермичният ефект на пептида може да бъде допълнителен бонус.
Работейки под ръководството на Гришин, открихме и инхибитор на P2X3 рецепторите. Това също се оказа пептид, който получи името PT1 и беше открит в отровата на паяк Alopecosa marikovskyi. Между другото, PT1 вече успешно премина лабораторни и предклинични тестове, така че след известно време може да се превърне в един от първите фундаментално нови аналгетици, които специфично инхибират рецепторите за „болка“. За третия от тези споменати рецептори, ASIC3, ние също открихме инхибитор: пептид Ugr 9-1; източникът беше отровата на морска анемона Urticina grebelnyi .
Имайте предвид, че естествените отрови често съдържат токсини с противоположен ефект, тоест вещества, които активират рецепторите за болка. От гледна точка на биологията на отровните животни това е разбираемо: те използват „болезнени“ токсини за отбранителни цели. Например в отровата на китайската тарантула Haplopelma schmidtiсъдържа най-силния активатор TRPV1 и от отровата на тексаската коралова змия Микрурус тенерПолучен е ASIC1a активатор. В днешно време вече сме се научили как да се възползваме от такива вещества: те се използват като молекулярни инструменти за „замразяване“ на рецепторите за болка в активирано състояние и изследване на тяхната структура (фиг. 3) [,]. От друга страна, откриването на полезни молекули в естествени отрови също е доста често срещано и няколко естествени токсини (или вещества, създадени от тях) сега се използват в медицината като лекарства. Това е мястото, където известната поговорка на средновековния алхимик Парацелз придобива специално значение: „Всичко е отрова и нищо не е без отрова; Само една доза прави отровата невидима.
Сензорните невронни рецептори представляват примамлива, но предизвикателна лекарствена цел. Лекарствата, ако имат добра селективност към тези рецептори, ще бъдат приети от потребителите с голяма радост, тъй като почти всички съвременни лекарства са ограничени в употреба поради странични ефекти. Работи се по търсенето на селективни лекарства, включително и у нас, и при благоприятно стечение на обстоятелствата такива скоро ще могат да се появят в аптеките. Дълъг живот без болка!
Тази работа беше подкрепена от Руската научна фондация (проект № 14-24-00118).
Литература
.
Palermo N.N., Brown H.K., Smith D.L. Селективно невротоксично действие на капсаицин върху гломерулни С-тип терминали в substantia gelatinosa на плъх // Brain Res. 1981. Т. 208. С. 506–510.
.
O'Neill J., Brock C., Olesen A.E. et al.
Физиология на болката
В тесния смисъл на думата болката е неприятно усещане, възникващо под действието на свръхсилни дразнители, предизвикващи структурни и функционални нарушения в организма. Разликата между болката и другите усещания е, че тя не информира мозъка за качеството на стимула, а показва, че стимулът е увреждащ. Друга особеност на сензорната система за болка е нейният най-сложен и мощен еферентен контрол.
Анализаторът на болката стартира няколко програми в централната нервна система за реакция на тялото към болка. Следователно болката има няколко компонента. Сетивният компонент на болката я характеризира като неприятно, болезнено усещане; афективен компонент – като силна негативна емоция; мотивационен компонент - като отрицателна биологична потребност, която предизвиква поведение на тялото, насочено към възстановяване. Моторният компонент на болката е представен от различни двигателни реакции: от безусловни флексионни рефлекси до двигателни програми на противоболково поведение. Вегетативният компонент характеризира дисфункцията на вътрешните органи и метаболизма при хронична болка. Когнитивният компонент е свързан със самооценката на болката, при която болката действа като страдание. Когато работят други системи, тези компоненти са слабо изразени.
Биологичната роля на болката се определя от няколко фактора. Болката действа като сигнал за заплаха или увреждане на телесните тъкани и ги предупреждава. Болката има когнитивна функция: чрез болката човек се научава да избягва възможните опасности от външната среда. Емоционалният компонент на болката изпълнява функцията на подсилване при формирането на условни рефлекси. Болката е фактор за мобилизиране на защитните и адаптивни реакции на организма при увреждане на неговите тъкани и органи.
Има два вида болка – соматична и висцерална. Соматичната болка се разделя на повърхностна и дълбока.Повърхностната болка може да бъде ранна (бърза, епична) и късна (бавна, протопатична).
Има три теории за болката.
1. Теорията за интензитета е предложена от Е. Дарвин и А. Голдщайнер. Според тази теория болката не е специфично чувство и няма свои специални рецептори. Това се случва, когато свръхсилни стимули действат върху рецепторите на петте известни сетивни органа. Конвергенцията и сумирането на импулси в гръбначния и главния мозък участват в образуването на болка.
2. Теорията за специфичността е формулирана от немския физиолог М. Фрей. Според тази теория болката е специфично чувство, което има собствен рецепторен апарат, аферентни влакна и мозъчни структури, които обработват информацията за болката. По-късно тази теория получи по-пълно експериментално и клинично потвърждение.
3. Съвременната теория за болката се основава предимно на теорията за специфичността. Доказано е съществуването на специфични рецептори за болка. В същото време съвременната теория на болката използва позицията за ролята на централната сумация и конвергенция в механизмите на болката. Най-големите постижения на съвременната теория за болката са разработването на механизми за централно възприятие на болката и задействането на противоболковата система на организма.
Рецептори за болка
Рецепторите за болка са свободните окончания на чувствителните миелинизирани нервни влакна Aδ и немиелинизираните влакна C. Те се намират в кожата, лигавиците, периоста, зъбите, мускулите, ставите, вътрешните органи и техните мембрани и кръвоносните съдове. Те не се намират в нервната тъкан на главния и гръбначния мозък. Тяхната най-голяма плътност се намира на границата на дентина и зъбния емайл.
Разграничават се следните основни типове рецептори за болка:
1. Механоноцицептори и механотермални ноцицептори на Aδ-влакна реагират на силни механични и термични стимули, провеждат бърза механична и термична болка, бързо се адаптират; разположени предимно в кожата, мускулите, ставите, периоста; техните аферентни неврони имат малки рецептивни полета.
2. Полисензорните ноцицептори на С-влакната реагират на механични, термични и химични стимули, провеждат късна, слабо локализирана болка и се адаптират бавно; техните аферентни неврони имат големи рецептивни полета.
Рецепторите за болка се стимулират от три вида стимули:
1. Механични дразнители, които създават налягане над 40 g/mm 2 при стискане, разтягане, огъване, усукване.
2. Топлинните дразнители могат да бъдат топлинни (> 45 0 C) и студени (< 15 0 С).
3. Химически дразнители, отделяни от увредени тъканни клетки, мастоцити, тромбоцити (К +, Н +, серотонин, ацетилхолин, хистамин), кръвна плазма (брадикинин, калидин) и окончанията на ноцицептивните неврони (субстанция Р). Някои от тях възбуждат ноцицепторите (К +, серотонин, хистамин, брадикинин, ADP), други ги сенсибилизират.
Свойства на рецепторите за болка: рецепторите за болка имат висок праг на възбуждане, което осигурява реакцията им само на екстремни стимули. Ноцицепторите на С-аферентите се адаптират слабо към дългодействащи стимули. Възможно е да се повиши чувствителността на рецепторите за болка - намаляване на прага на тяхното дразнене при многократно или продължително стимулиране, което се нарича хипералгезия. В този случай ноцицепторите са способни да реагират на стимули с подпрагова величина, както и да бъдат възбудени от стимули от други модалности.
Пътища на чувствителност към болка
Невроните, които възприемат болковите импулси. От рецепторите за болка на тялото, шията и крайниците, Aδ- и C-влакната на първите сетивни неврони (телата им са разположени в гръбначните ганглии) отиват като част от гръбначните нерви и навлизат през дорзалните коренчета в гръбначния мозък , където се разклоняват в дорзалните колони и образуват синаптични връзки директно или чрез интерневрони с втори сензорни неврони, чиито дълги аксони са част от спиноталамичните пътища. В същото време те възбуждат два вида неврони: някои неврони се активират само от болезнени стимули, други - конвергентни неврони - също се възбуждат от неболезнени стимули. Вторите неврони на чувствителност към болка са предимно част от страничните спиноталамични пътища, които провеждат повечето от болковите импулси. На нивото на гръбначния мозък аксоните на тези неврони се преместват в страната, противоположна на стимулацията; в мозъчния ствол те достигат до таламуса и образуват синапси върху невроните на неговите ядра. Част от болковите импулси на първите аферентни неврони се прехвърлят чрез интернейрони към двигателните неврони на мускулите флексори и участват във формирането на защитни болкови рефлекси. В латералния спиноталамичен тракт се разграничават еволюционно по-младият неоспиноталамичен тракт и древният палеоспиноталамичен тракт.
Неоспиноталамичният път провежда сигнали за болка по Aδ влакна главно до специфични сензорни (вентрално задни) ядра на таламуса, които имат добра топографска проекция към периферията на тялото. В допълнение, малка част от импулсите навлизат в ретикуларната формация на багажника и след това в неспецифичните ядра на таламуса. Предаването на възбуждане в синапсите на този път се осъществява с помощта на бързодействащия предавател глутамат. От специфичните ядра на таламуса сигналите за болка се предават предимно към сензорната кора на мозъчните полукълба. Тези характеристики формират основната функция на неоспиноталамичния път - провеждане на "бърза" болка и възприемането й с висока степен на локализация.
Палеоспиноталамичният път провежда сигнали за болка по С-влакна главно към неспецифичните ядра на таламуса директно или след превключване в невроните на ретикуларната формация на мозъчния ствол. Предаването на възбуждане в синапсите по този път става по-бавно. Медиаторът е субстанция Р. От неспецифичните ядра импулсите навлизат в сетивните и други части на мозъчната кора. Малка част от импулса навлиза и в специфични ядра на таламуса. По принцип влакната на този път завършват на неврони на 1) неспецифични ядра на таламуса; 2) ретикуларна формация; 3) централно сиво вещество; 4) синьо петно; 5) хипоталамус. „Късната“, слабо локализирана болка се предава по палеоспиноталамичния път и се формират афективни и мотивационни прояви на чувствителност към болка.
В допълнение, чувствителността към болка се осъществява частично чрез други възходящи пътища: предните спиноталамични, тънки и клиновидни пътища.
Горните пътища провеждат и други видове чувствителност: температурна и тактилна.
Ролята на мозъчната кора в усещането за болка
Пълното сетивно възприемане на болката от тялото без участието на кората на главния мозък е невъзможно.
Основното проекционно поле на анализатора на болката се намира в соматосензорния кортекс на задния централен гирус. Осигурява възприемане на „бърза“ болка и идентифициране на нейното местоположение върху тялото. За по-точно идентифициране на локализацията на болката процесът задължително включва неврони на моторната кора на предния централен гирус.
Вторичното проекционно поле се намира в соматосензорния кортекс на границата на пресечната точка на централната бразда с горния ръб на темпоралния лоб. Невроните на това поле имат двустранни връзки с ядрата на таламуса, което позволява на това поле селективно да филтрира болезнените възбуди, преминаващи през таламуса. А това от своя страна позволява това поле да бъде включено в процеси, свързани с извличане на енграмата на необходимия поведенчески акт от паметта, внедряването му в дейността на ефекторите и оценка на качеството на постигнатия полезен резултат. Моторните компоненти на болковото поведение се формират в съвместната дейност на моторния и премоторния кортекс, базалните ганглии и малкия мозък.
Фронталната кора играе важна роля при възприемането на болката. Осигурява самооценка на болката (нейния когнитивен компонент) и формиране на целенасочено болково поведение.
Лимбичната система (cingulate gyrus, hipocampus, dentate gyrus, амигдала комплекс на темпоралния лоб) получава информация за болка от предните ядра на таламуса и формира емоционалния компонент на болката, задейства автономни, соматични и поведенчески реакции, които осигуряват адаптивни реакции към болезнен стимул.
Някои видове болка
Има болки, които са назовани проекцияили фантом. Възникването им се основава на закона за проекцията на болката: без значение коя част от аферентния път е раздразнена, болката се усеща в областта на рецепторите на този сензорен път. Според съвременните данни във формирането на този вид болка участват всички звена на болковата сетивност.
Има и т.нар отразениболка: когато се усеща болка не само в засегнатия орган, но и в съответния дерматом на тялото. Участъците от повърхността на тялото на съответния дерматом, където възниква усещането за болка, се наричат Зони на Захарьин-Гед. Появата на посочената болка се дължи на факта, че невроните, носещи болкови импулси от рецепторите на засегнатия орган и кожата на съответния дерматом, се събират в същия неврон на спиноталамичния тракт. Дразненето на този неврон от рецепторите на засегнатия орган в съответствие със закона за проекцията на болката води до факта, че болката се усеща и в областта на кожните рецептори.
Антиноцицептивна система
Противоболковата система се състои от четири нива: спинално, мозъчно-стволово, хипоталамусно и кортикално.
1. Спинално ниво на антиноцицептивната система. Неговият важен компонент е „контролът на вратата“ на гръбначния мозък, чиято концепция има следните основни принципи: предаването на болкови нервни импулси от първите неврони към невроните на спиноталамичния тракт (втори неврони) в задните колони на гръбначният мозък се модулира от механизма на гръбначната врата - инхибиторни неврони, разположени в желатиновата субстанция на гръбначния мозък. Разклонените аксони на различни сензорни пътища завършват на тези неврони. На свой ред невроните на желатиновата субстанция упражняват пресинаптично инхибиране в местата на превключване на първия и втория неврон на болката и други сензорни пътища. Някои неврони са конвергентни: върху тях невроните образуват синапси не само от рецептори за болка, но и от други рецептори. Контролът на спиналния портал се регулира от съотношението на импулсите, пристигащи по протежение на аферентни влакна с голям диаметър (неболкова чувствителност) и малък диаметър (болкова чувствителност). Интензивният поток от импулси по влакна с голям диаметър ограничава предаването на сигнали за болка към невроните на спиноталамичните пътища (затваря "портата"). Напротив, интензивен поток от болкови импулси по протежение на първия аферентен неврон, инхибирайки инхибиторните интерневрони, улеснява предаването на сигнали за болка към невроните на спиноталамичните пътища (отваря "портата"). Механизмът на гръбначния стълб е под постоянното влияние на нервните импулси от структурите на мозъчния ствол, които се предават по низходящи пътища както към невроните на желатиновата субстанция, така и към невроните на спиноталамичния тракт.
2. Ниво на мозъчния ствол на антиноцицептивната система. Стволовите структури на аналгетичната система включват, първо, централното сиво вещество и raphe ядрата, образуващи един функционален блок, и второ, магноцелуларните и парагиантните клетъчни ядра на ретикуларната формация и locus coeruleus. Първи комплексблокира преминаването на болкови импулси на нивото на релейните неврони на ядрата на дорзалните рога на гръбначния мозък, както и на релейните неврони на сензорните ядра на тригеминалния нерв, образувайки възходящи пътища на чувствителност към болка. Вторият комплекс възбужда почти цялата антиноцицептивна система (виж фиг. 1).
3. Хипоталамусното ниво на антиноцицептивната система, от една страна, функционира независимо, а от друга, действа като настройка, която контролира и регулира антиноцицептивните механизми на ниво ствол поради връзките между хипоталамичните неврони с различна ядрена принадлежност и различни неврохимични специфики. Сред тях са идентифицирани неврони, в чиито окончания се отделят енкефалини, β-ендорфин, норепинефрин и допамин (виж фиг. 2).
4. Кортикално ниво на антиноцицептивната система. Соматосензорната област на мозъчната кора обединява и контролира активността на антиноцицептивните структури на различни нива. В този случай най-важната роля в активирането гръбначни и стволови структурииграе вторичната сензорна област. Невроните му образуват най-голям брой влакна за низходящ контрол на чувствителността към болка, насочващи се към дорзалните рога на гръбначния мозък и ядрата на мозъчния ствол. Вторичната сензорна кора модифицира активността на стволовия комплекс на антиноцицептивната система. В допълнение, соматосензорните полета на кората на главния мозък контролират провеждането на аферентни болкови импулси през таламуса. В допълнение към таламуса, кората на главния мозък регулира преминаването на болковите импулси в хипоталамуса, лимбичната система, ретикуларната формация и гръбначния мозък. Водещата роля в осигуряването на кортико-хипоталамични влияния се възлага на невроните на фронталния кортекс.
Медиатори на антиноцицептивната система
Медиаторите на аналгетичната система включват пептиди, които се образуват в мозъка, аденохипофизата, надбъбречната медула, стомашно-чревния тракт, плацентата от неактивни прекурсори Сега опиатните медиатори на антиноцицептивната система включват: 1) ά-, β-, γ-ендорфини; 2) енкефалини; 3) динорфини. Тези медиатори действат върху три типа опиатни рецептори: μ-, δ-, κ-рецептори. Най-селективният стимулатор на μ-рецепторите са ендорфините, δ-рецепторите са енкефалините, а κ-рецепторите са динорфините. Плътността на μ- и κ-рецепторите е висока в мозъчната кора и гръбначния мозък и средна в мозъчния ствол; плътността на δ-рецепторите е средна в кората на главния мозък и гръбначния мозък, ниска в мозъчния ствол. Опиоидните пептиди инхибират действието на веществата, които причиняват болка на ниво ноцицептори, намаляват възбудимостта и проводимостта на болковите импулси и инхибират предизвиканата реакция на невроните, разположени във веригите, които предават болковите импулси. Тези пептиди достигат до невроните на сензорната система за болка с кръв и цереброспинална течност. Опиоидните медиатори се освобождават в синаптичните окончания на невроните на аналгетичната система. Аналгетичният ефект на ендорфините е висок в главния и гръбначния мозък, ефектът на енкефалините в тези структури е среден, ефектът на динорфините в мозъка е слаб, а в гръбначния мозък е висок.
Фиг. 1. Взаимодействие на основните елементи на аналгетичната система от първо ниво: мозъчен ствол - гръбначен мозък. (отворените кръгове са възбуждащи неврони, черните кръгове са инхибиращи).
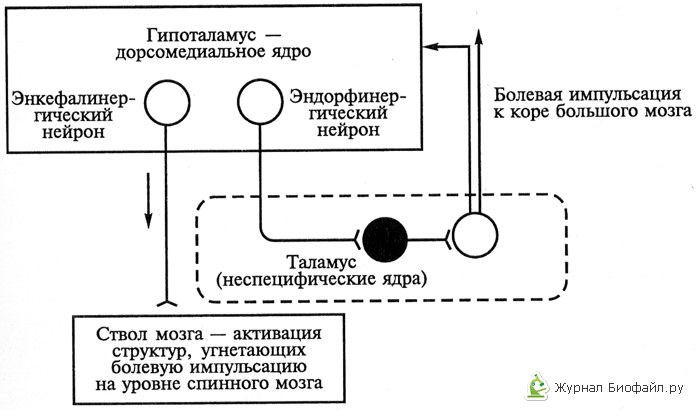
Фиг.2. Механизмът на работа на системата за облекчаване на болката от второ ниво на тялото (хипоталамус - таламус - мозъчен ствол) с помощта на опиоиди.
Светлите кръгове са възбуждащи неврони, черните кръгове са инхибиращи.
Тежестта на болката не се определя само от силата на екзогенната или ендогенната болка. Това до голяма степен зависи от съотношението на активността на ноцицептивните и антиноцицептивните части на болковата система, което има адаптивно значение.
Повърхностните тъкани са снабдени с нервни окончания на различни аферентни влакна ( J.Erlanger, Г. С. Гасер, 1924). Най-дебелите, миелинизирани Ab влакна имат тактилна чувствителност. Те се възбуждат от неболезнено докосване и движение. Тези окончания могат да служат като мултимодални неспецифични рецептори за болка само при патологични състояния, например поради повишаване на тяхната чувствителност (сенсибилизация) от възпалителни медиатори. Слабото стимулиране на мултимодални неспецифични тактилни рецептори води до чувството сърбеж. Прагът на тяхната възбудимост се понижава от хистамин и серотонин ( Г. Щютген, 1981).
Специфични първични рецептори за болка (ноцицептори) са два други вида нервни окончания - тънки миелинизирани Ad-терминали и тънки немиелинизирани С-влакна, които са филогенетично по-примитивни. И двата типа терминали присъстват както в повърхностните тъкани, така и във вътрешните органи. Някои области на тялото, като роговицата, се инервират само от Ad и C аференти. Ноцицепторите дават усещане за болка в отговор на различни интензивни стимули - механично въздействие, термичен сигнал (обикновено с температура над 45-47 0 C), дразнещи химикали, например киселини. Исхемията винаги причинява болка, защото провокира ацидоза. Мускулният спазъм може да предизвика дразнене на болковите окончания поради относителната хипоксия и исхемия, които причинява, както и поради директното механично изместване на ноцицепторите.
Бавната, протопатична болка се провежда през С-влакна със скорост 0,5-2 m/sec, а епикритичната болка се провежда през миелинизирани, бързопроводими Ad-влакна, осигуряващи скорост на проводимост от 6 до 30 m/sec. С изключение на кожата, където по данни А. Г. Бухтиярова(1966), има най-малко 100-200 рецептора за болка на 1 cm 2, лигавиците и роговицата, периостът е изобилно снабден с рецептори за болка от двата вида (както всеки футболист, който получава удар по предно-вътрешната повърхност на пищяла по време на борба), както и съдовите стени, ставите, церебралните синуси и париеталните слоеве на серозните мембрани.
Във висцералните слоеве на тези мембрани и вътрешните органи има много по-малко рецептори за болка. Освен това в паренхима на вътрешните органи има изключително С-влакна с протопатична чувствителност, достигащи до гръбначния мозък като част от автономните нерви. Следователно висцералната болка е по-трудна за локализиране от повърхностната болка. В допълнение, локализирането на висцералната болка зависи от феномена на „отнесената болка“, чиито механизми са обсъдени по-долу. Париеталният перитонеум, плеврата, перикардът, капсулите на ретроперитонеалните органи и част от мезентериума имат не само бавни протопатични С-влакна, но и бързи епикритични Ad влакна, свързани с гръбначния мозък чрез гръбначни нерви. Следователно болката от тяхното дразнене и увреждане е много по-остра и по-ясно локализирана. Дори в преданестезиологичната ера хирурзите забелязаха, че чревните разрези са по-малко болезнени от разрезите на париеталния перитонеум. Болката по време на неврохирургични операции е максимална в момента на дисекция на менингите, докато в същото време мозъчната кора има много незначителна и строго локална чувствителност към болка. Като цяло, такъв често срещан симптом като главоболие, почти винаги се свързва с дразнене на рецепторите за болка извън самата мозъчна тъкан. Екстракраниалната причина за главоболие може да бъде процеси, локализирани в синусите на костите на главата, спазъм на цилиарния и други очни мускули, тонично напрежение на мускулите на шията и скалпа. Интракраниалните причини за главоболие са преди всичко дразнене на ноцицепторите в менингите. При менингит силното главоболие обхваща цялата глава. Много сериозно главоболие се причинява от дразнене на ноцицепторите в церебралните синуси и артерии, особено в средната церебрална артерия. Дори незначителни загуби на цереброспинална течност (около 20 ml) могат да провокират главоболие, особено в изправено положение на тялото, тъй като плаваемостта на мозъка се променя и когато хидравличната възглавница намалява, рецепторите за болка на мембраните му се дразнят. От друга страна, излишъкът на гръбначно-мозъчната течност и нарушаването на изтичането й при хидроцефалия, мозъчен оток, подуване при вътреклетъчна хиперхидратация, конгестия на съдовете на менингите, причинена от цитокини при инфекции, локални обемни процеси също провокират „най-честото оплакване“ - главоболие , така че как това увеличава механичния ефект върху рецепторите за болка на структурите около самия мозък. Общият принцип на локализиране на главоболието е такъв, че тилната болка често отразява дразненето на ноцицепторите на съдовете и менингите под тенториума, а супралаталните дразнители и стимулацията на горната повърхност на самата палатка се проявяват чрез фронто-париетална болка. „Главоболието от махмурлук“, познато на много голяма част от човечеството, има сложна патогенеза, включително предизвикана от алкохол конгестия на менингите и вътреклетъчна хиперхидратация. Патофизиологията на някои форми на главоболие, тясно свързани с хуморалните медиатори на болковата и антиболковата системи и с механизмите на провеждане на тези системи, по-специално мигрена, се обсъжда отделно по-долу.
Паренхимът на далака, бъбреците, черния дроб и белия дроб е напълно лишен от ноцицептори. Но бронхите, жлъчните пътища, капсулите и съдовете на тези органи са богато снабдени с тях. Дори големи абсцеси на черния дроб или белия дроб могат да бъдат почти безболезнени. Плевритът или холангитът обаче понякога причиняват сериозна болка, без да са тежки сами по себе си. Рецепторите за висцерална болка също се различават по това, че развиват относително слаб отговор на строго локално увреждане на орган, например хирургически разрез. Въпреки това, при дифузно засягане на тъканта в промяната (на фона на исхемия, под действието на литични ензими и дразнещи химикали, със спазми и хиперекстензия на кухи органи), тяхната чувствителност под въздействието на възпалителни медиатори бързо се увеличава и се излъчват силни импулси от тях.
Рецепторите за болка претендират за уникална позиция в човешкото тяло. Това е единственият тип сензорен рецептор, който не подлежи на никаква адаптация или десенсибилизация под въздействието на непрекъснат или повтарящ се сигнал. Ноцирецепторите не повишават своя праг на възбудимост, както правят други, например сензорите за студ. Следователно рецепторът не "свиква" с болката. Освен това в ноцирецептивните нервни окончания се получава точно обратното явление - сенсибилизиране на болковите рецептори чрез сигнал. При възпаление, увреждане на тъкани (особено вътрешни органи) и повтарящи се и продължителни болезнени стимули, прагът на ноцицепторната възбудимост намалява. Дори и най-лекото докосване до изгорената повърхност е изключително болезнено. Това явление се нарича първична хипералгезия. Палпацията на вътрешните органи, дори и да е интензивна, не причинява болка, ако няма възпаление. По време на възпаление обаче чувствителността на тихите вътрешни ноцицептори се увеличава толкова много, че лекарят регистрира симптоми на болка. Потупването на областта на бъбреците, безболезнено при липса на увреждане, води до болка, ако бъбречните ноцицептори са сенсибилизирани от възпалителни медиатори (положителен симптом на Пастернацки). Лесно е да се отбележи, че ако настъпи адаптация на рецепторите за болка, всички хронични деструктивни процеси биха били безболезнени и болката би загубила функцията си на сигнал, което според израза И. П. Павлова, „насърчава ви да изхвърлите това, което застрашава жизнения процес.“
Наричайки сензорите за болка рецептори, трябва да подчертаем, че прилагането на този термин към тях е условно - все пак това са свободни нервни окончания, лишени от специални рецепторни устройства.
Неврохимичните механизми на ноцицепторното дразнене са добре проучени. Основният им стимулатор е брадикининът. В отговор на увреждане на клетки в близост до ноцирецептора, този медиатор, както и простагландини, левкотриени и калиеви и водородни йони, се освобождават. Простагландините и левкотриените сенсибилизират ноцицепторите към кинините, а калият и водородът улесняват тяхната деполяризация и възникването на електрически аферентен болков сигнал в тях. Възбуждането се разпространява не само аферентно, но и антидромно към съседните разклонителни терминали. Там води до секреция на субстанция Р. Този невропептид, който вече беше споменат, причинява хиперемия, оток, дегранулация на мастоцитите и тромбоцитите около терминала по паракринен начин. Освободените в този случай хистамин, серотонин и простагландини сенсибилизират ноцицепторите, а химазата и триптазата на мастните клетки засилват производството на техния директен агонист брадикинин. Следователно, когато са увредени, ноцирецепторите действат едновременно като сензори и като паракринни провокатори на възпаление. В близост до ноцицепторите, като правило, има симпатични норадренергични постганглионарни нервни окончания, които са способни да модулират чувствителността на ноцицепторите. При увреждания на периферни нерви, т.нар каузалгия- патологично повишена чувствителност на ноцицепторите в областта, инервирана от увредения нерв, придружена от пареща болка и дори признаци на възпаление без видимо локално увреждане. Механизмът на каузалгията е свързан с хипералгичния ефект на симпатиковите нерви, по-специално на секретирания от тях норепинефрин, върху състоянието на рецепторите за болка. Възможно е субстанция Р и други невропептиди да се секретират от симпатиковите нерви, което причинява възпалителни симптоми. Феноменът на каузалгията е в пълния смисъл неврогенно възпаление, въпреки че се причинява не от нервен, а от паракринен метод (виж също по-горе, за ролята на нервната регулация при възпаление).
Както първоначално беше предложено У. КенънИ А. Розенблут(1951) паракринната безимпулсна невропептидергична активност на нервните окончания в тъканите е истинската основа на феномена, който повече от 100 години от Ф. Магенди(1824) до Ел Ей Орбели(1935) и ПО дяволите. Сперански, (1937), нар нервна трофика.
Дата на добавяне: 2015-05-19 | Видяно: 985 | Нарушаване на авторски права
| | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | | |